
| Crop Knowledge Master | ||
|
Brevipalpus phoenicis (Geijskes) |
|
| Red and Black Flat Mite | ||
| Hosts | Distribution | Damage | Biology | Behavior | Management | Reference | |
Authors
Jayma L. Martin Kessing, Educational Specialist
Ronald F.L. Mau, Extension Entomologist
Department of Entomology
Honolulu, Hawaii
This mite has an extensive host range and may cause economic damage, depending on the host. Pritchard and Baker (1958) list over 65 hosts. In Hawaii, the red and black flat mite has been reported on anthurium, banana, Hemigraphis, lemon, macadamia, orchid, papaya and passion fruit. In other parts of the world it is common on tea and Citrus.
This is a tropical-subtropical species that has been accidentally transported by man to many areas of North America where it survives in greenhouses. This mite was first found in Hawaii on Oahu in 1955 and has subsequently been reported on Kauai, the Big Island and Maui.
In Hawaii, the red and black flat mite is abundant in areas between sea level and 1,000 feet, scarce between 1,000 and 2,500 feet and has not been recorded in areas above 2,500 feet in elevation (Haramoto, 1969). It is usually not considered to be a pest of economic importance above 1,000 feet (Haramoto, 1969).
Feeding by this mite devitalizes the plant and causes the collapse of the inner leaf tissue. Old damage is characterized by browning of the damaged leaf surface. In addition, some hosts exhibit deformed leaves.
On papaya plants, this mite usually feeds on the trunk six to ten inches below the attachment of the bottom whorl of leaves (Haramoto, 1969). The mites move upward on the trunk and outward onto the leaves and fruit as the population density increases. Many feeding punctures occur close together, and the affected areas coalesce to form large continuous callus-like, light brown, scaly and/or scabby areas (Haramoto, 1969). Feeding damage is pronounced on young papayas and injured areas become sunken because they grow at a different rate than the unaffected tissues. In heavy infestations, the papaya stem becomes tan colored and suberized prematurely and makes a spindly growth (Haramoto, 1969).
On citrus, this mite has mean associated as the cause of swellings on citrus stems ("Brevipalpus gall"). Also leaves abscise prematurely or exhibit foliar chlorosis ("phoenicis blotch") (Knorr, et. al., 1960).
Populations are primarily composed of females; males compose less than 1% of the population in Hawaii (Haramoto, 1969). Reproduction primarily occurs through unfertilized parthenogenesis. Eggs produce only females. Because of the size of these insects it is very difficult to see the different stages without the use of a microscope. Refer to Haramoto (1969) and Nagesha et al. (1974) for detailed descriptions.
The duration of different life stages varies with temperature and humidity. The duration from newly laid egg to adult requires minimum of 18.6 days at 86û F and a maximum of 48.8 days at 68û F under laboratory conditions (Haramoto, 1969). The mite cannot complete its life cycle at humidities below 30%, nor at average temperatures above 86û F and below 68û F (Haramoto, 1969).
Generations are continuous in Hawaii, often overlapping. Based on laboratory studies there are at least 10 generations per year (Haramoto, 1969).
On Oroxylum indicum the life cycle was completed in 20.02 days at 70û F and 29.66 days at 79û F (Lal, 1979). In the same study by Lal (1979), the life cycle duration on Clerodendron siphonanthus took 28.34 and 20.20 days at 70û F and 79û F, respectively.
EGGS
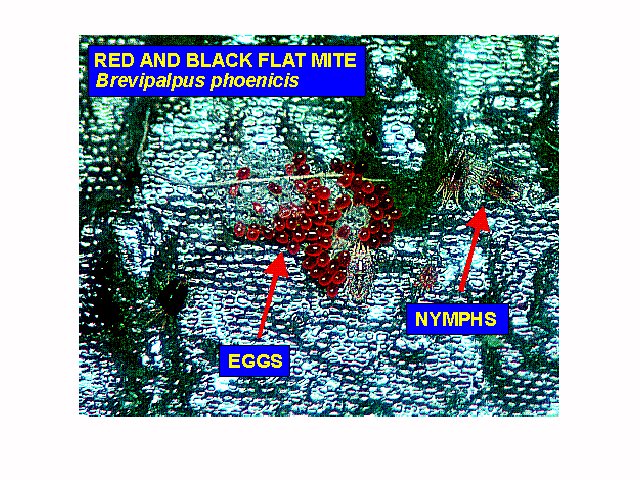
Eggs are deposited in cracks, crevices and other protected areas on the plant surface. Although each egg is laid singly, they often occur clustered together because females will use an egg laying site several times (Haramoto, 1969). These clusters of bright reddish orange eggs are more easily seen with the naked eye than any other life stage (Haramoto, 1969).
Eggs are elliptical and about 1/250 inch long by 7/2500 inch wide (Haramoto, 1969) and slightly broadened at one end. When first laid they are light orange, soft and very sticky. At this time they readily adhere to any surface. Eggs have a stipe, a tail-like projection, that extends from the slightly pointed end that came out of the female mite last. This stipe often breaks off if the egg is handled. A day before hatching, the eggs become opaque white and the red eyes of the larvae are visible within (Haramoto, 1969).
Hatching occurs in minimum of 8 days at 86û F with 65 to 70 percent humidity and a maximum of 24.8 days at 68û F with 85 to 90 percent humidity under laboratory conditions (Haramoto, 1969). Eggs do not hatch in constant temperatures below 68û F and above 86û F regardless of humidity (Haramoto, 1969).
LARVAE
Larvae are 6 legged, bright orange-red when newly emerged and average about 14/2500 inch long by 9/2500 inch wide (Haramoto, 1969). When fully grown they are opaque orange and 17/2500 inch long by 11/2500 inch wide (Haramoto, 1969).
NYMPHS
There are two nymphal stages, the protonymph and the deutonymph. The protonymph is larger than the larva (about 23/2500 inch long by 14/2500 inch wide) and has 8 legs. The outer shell is transparent, light green, orange, black and yellow patches may be seen within the body. The deutonymph is similar in appearance to the protonymph except for having an extra pair of legs, two additional setae (hairs) and being slightly larger (averaging 29/2500 inch long by 16/2500 inch wide) (Haramoto, 1969).
Duration from egg hatch to adult required a minimum of 10.6 days at 86û F and a maximum of 27.3 days at 68û F under laboratory conditions (Haramoto, 1969). The optimum development duration of 19.8 days occurred at 77û F with 70 percent humidity (Haramoto, 1969). Temperatures above 86û F and below 68û F for prolonged periods are fatal to immature stages.
ADULTS
The adult female is very small, approximately 3/250 inch long by 16/2500 inch wide (Haramoto, 1969). The body is elliptical, flat, light to dark green or reddish orange. There are four legs extending forward and 4 legs extending behind. A black mark in the shape of an "H" becomes visible when these mites are reared in temperatures between 68û F and 77û F, while this black spot is not present at 86û F (Haramoto, 1969).
The adult male is flat, reddish and more wedge shaped than the female. Males do not have black markings (Haramoto, 1969).
On average, adults lived for a maximum of 47 days at 68û F and a minimum of 7.5 days at 86û with a relative humidity of 85 to 90 percent (Haramoto, 1969).
During the pre-oviposition period, females feed for 4 to 12 days (average 7.74 days) on guava (Nagesha Chandra and Channabasavanna, 1974) and 2.3 to 6.5 days on papaya (Haramoto, 1969). At 77û F each female laid from one to four eggs per day (Haramoto, 1969) for 9 to 35 days (Nagesha Chandra and Channabasavanna, 1974). The number of eggs laid by each female varies with temperature if the relative humidity is held constant (Haramoto, 1969). The oviposition process itself takes 12 to 30 minutes (Zaher, et. al., 1971). Females live for 2 to 22 days (average 10.4 days) after she has stopped laying eggs on guava (Nagesha Chandra and Channabasavanna, 1974).
Larval, protonymph and deutonymph stages have active periods during which they feed, grow and disperse followed by an inactive stage in which they transform into the next stage of development. Feeding, growth and dispersal activity are similar in all stages becoming more pronounced with the passing of each stage (Haramoto, 1969). During the inactive stage this mite assumes a characteristic position with its feeding stylets penetrating the plant tissue and legs straightened outward. There is no feeding during this period and the stylets anchor the mite to the plant (Haramoto, 1969).
The red and black flat mite feeds throughout the day and night. Feeding is especially intense in temperatures between 77û and 86û F and high relative humidity (Haramoto, 1969). They generally feed on the stems of plants, but will feed on fruits when population densities are high (Haramoto, 1969). Females start feeding soon after emergence and must do so before laying eggs. Females usually remain near the area in which they hatched if food is ample and suitable.
NON-CHEMICAL CONTROL
The abundance of host plants, warm and humid climate and few natural enemies are favorable conditions for high population densities of this mite (Haramoto, 1969). Conditions contrary to these do not support high population densities and favor control by natural enemies (Haramoto, 1969).
Biological Control -- Predators
At least four predators of the red and black flat mite are known in Hawaii: three mites: Phytoseiulus macropilis (Banks), Amblyseius largoensis (Muma), Mexecheles hawaiiensis (Baker); and the beetle, Sticholotis punctata Crotch (Haramoto, 1969). The predatory mites, P. macropilis and A. largoensis feed on the eggs the red and black mite, but are not suspected to live and reproduce exclusively on them (Haramoto, 1969). The larvae, nymphs and adults of M. hawaiiensis attack all active stages of the red and black flat mite and may survive solely on them. Accounting for 87%, M. hawaiiensis is the most abundant predator of this pest in Hawaii. S. punctata was introduced from China and Japan to control scales, it can noticeably control the red and black mite and feeds on all stages (Haramoto, 1969).
The above mentioned predators generally do not provide economic control because their predatory activity becomes apparent when the prey population density is very high and severe plant damage has already occurred. Therefore, other agents must be introduced for economic control (Haramoto, 1969).
CHEMICAL CONTROL
Because this mite reproduces parthenogenically, as do most other false spider mites, they do not have the genetic variability and mixing of most mite species that reproduce sexually. Therefore, resistance to pesticides is less likely to be selected out from a population (Haramoto, 1969) and is less of a problem. Although the red and black flat mite is resistant to certain organophosphates, it is susceptible to most acaricides. However, resistance may develop if selection pressure is high due to frequent pesticide applications.
Chemical sprays should be applied to affected plants at 2-3 week intervals if the infestation is severe (Hill, 1983). Any of the registered miticides are usually effective. Wettable sulfur should not be used in localities where relative humidity is abnormally low.
Denmark, H. A. 1975. A False Spider Mite, Brevipalpus phoenicis (Geijskes), Damage to Aphelandra. Fla. Dept. Agr. & Consumer Serv., Division of Plant Industry, Entomology Circular No. 54.
Haramoto, F. H. 1968. Biology and Control of Brevipalpus phoenicis (Geijskes) (Acarina: Tenuipalpidae). Hawaii Agric. Exp. Sta. Tech. Bull. No. 68: 1-63.
Hill, D. S. 1983. Brevipalpus phoenicis (Geijskes). pp. 503. In Agricultural Insect Pests of the Tropics and Their Control, 2nd Edition. Cambridge University Press. 746 pages.
Knorr, L. C. and H. A. Denmark. 1970. Injury to Citrus by the Mite Brevipalpus phoenicis. J. Econ. Entomol. 63(6): 1996-1998.
Knorr, L. C., B. N. Webster and G. Malaguti. 1960. Injuries to Citrus Attributed to Brevipalpus Mites, including Brevipalpus Gall, a Newly Reported Disorder in Sour-Orange Seedlings. FAO (Food Agr. Organ. U.N.) Plant Prot. Bull. 8:141-148.
Lal, L. 1979. Biology of Brevipalpus phoenicis (Geijskes) (Tenuipalpidae: Acarina). Acarologia. 20(1): 97-101.
Nagesha Chandra, B. K. and G. P. Channabasavanna. 1974. Biology of Guava Scarlet Mite, Brevipalpus phoenicis (Geijskes) (Acarina: Tenuipalpidae). Proc. of the 4th International Congress of Acarology. 167-176.
Pritchard, A. E. and E. W. Baker. 1958. The False Spider Mites (Acarina: Tenuipalpidae). Univ. Calif. Publ. Ent. 14(3): 175-274, 51 fig.
Zaher, M. A., A. K. Wafa and A. A. Yousef. 1971. Biology of Brevipalpus phoenicis (Geijskes), in Egypt. Soc. Entomol. Egypte Bull. 5: 177-183.
AUG/1992.
B-PHOENI
{kind=link}
{kind=link}
{kind=link}